




[ad_1]
Disclaimer: Early release articles are not considered as final versions. Any changes will be reflected in the online version in the month the article is officially released.
Author affiliation: Author affiliations: Institute of Antibiotics, Huashan Hospital, Fudan University, Shanghai, China (Y. Shao, M. Wang, Q. Guo); Key Laboratory of Clinical Pharmacology of Antibiotics, National Heath Commission of the People’s Republic of China, Shanghai (Y. Shao, M. Wang, Q. Guo); Minhang Hospital, Fudan University, Shanghai (M. Chen); Children’s Hospital of Fudan University, Shanghai (J. Cai, M. Zeng); University of Pittsburgh School of Medicine, Pittsburgh, Pennsylvania, USA (Y. Doi); Fujita Health University School of Medicine, Toyoake, Japan (Y. Doi); Shanghai Municipal Center for Disease Control and Prevention, Shanghai (M. Chen)
Invasive meningococcal disease (IMD) is a severe infection caused by Neisseria meningitidis. Early empiric treatment with penicillin or third-generation cephalosporins (3GCs) including ceftriaxone and cefotaxime, is crucial (1). Over the past 2 decades, the spread of the hyperinvasive and quinolone-resistant N. meningitidis clone ChinaCC4821-R1-C/B (sublineage L44.1, mainly serogroup C) within clonal complex (CC) 4821 has resulted in ≈70% fluoroquinolone resistance among meningococci in China (2). More recently, penicillin nonsusceptiblity has increased rapidly among meningococci in China (3–5). We recently reported acquisition of penicillin and cefotaxime resistance by CC4821 (3).
The penA gene (around 1,746 bp, NEIS1753) encodes penicillin binding protein 2 (PBP2), a 2-domain protein (6). A 402-bp region (nucleotides 1321–1722, amino acids 441–574) is commonly used to determine penA alleles in meningococci. Five well-characterized alterations in the C-terminal catalytic transpeptidase of PBP2 (PBP2-TPase) are primarily responsible for penicillin nonsusceptibility in meningococci (7). A series of additional mutations in PBP2-TPase contribute to ceftriaxone resistance in the globally disseminated N. gonorrhoeae FC428 clone (8,9). Three cefotaxime-resistant meningococci isolates were reported in China during 2017–2019 (3). The case reported in this article involves, 1 of those 3 isolates, Nm507, serogroup C quinolone-resistant sequence type (ST) 4821 (C: P1.21–2,9:F3–3:ST4821, L44.1), which led to a rare fulminant case of IMD (Appendix). This study was approved by the Institutional Review Board of Children’s Hospital of Fudan University (approval no. 2023–111).
Figure 1

Figure 1. Epidemiologic and molecular characterizations of penA795-bearing Neisseria meningitidis, N. gonorrhoeae, and Neisseria commensals in study of cefotaxime-resistant N. meningitidissequence type…
A 2-year-old child was initially infected with influenza A virus and subsequently developed purpura fulminans, progressing to septic shock and disseminated intravascular coagulation. Mechanical ventilation, fluid resuscitation, and vasoactive drugs were administered to stabilize his vital signs. After 1 dose of penicillin, antimicrobial therapy was changed to ceftriaxone for 14 days (Appendix Figure 1). The child recovered fully. Previous influenza A virus infection is hypothesized to increase the risk for IMD because it disrupts the nasopharyngeal epithelium and normal flora (10).
We isolated 3 N. meningitidis isolates from the patient: Nm507 (blood), Nm508 (cerebrospinal fluid), and Nm509 (nasopharynx). Antimicrobial susceptibility testing showed the 3 N. meningitidis isolates were resistant to penicillin (MICs 0.75 μg/mL), cefotaxime (MICs 0.25 μg/mL), ciprofloxacin (MICs 0.5 μg/mL), and trimethoprim/sulfamethoxazole (MICs >2/38 μg/mL) and had reduced susceptibility to ceftriaxone (MICs 0.125 μg/mL) and meropenem (MICs 0.047 μg/mL). The 3 isolates were genetically identical and harbored penA795, which contained 12 mutations associated with penicillin and 3GCs resistance in the PBP2-TPase. These mutations included 5 classic penicillin-resistance–associated mutations (F504L, A510V, I515V, H541N, and I566V) (7) and the A549T mutation, which contributes to penicillin nonsusceptibility (3). In addition, they contained 6 mutations found in N. gonorrhoeae strain FC428 associated with resistance to 3GCs (A311V, I312M, V316T, T483S, N512Y, and G545S) (8,9), without any other determinants conferring resistance to 3GCs. Natural transformants, obtained from previous research (3), harbored mosaic penA, and the segments containing penA795 were transferred from 3 commensal Neisseria species to penicillin- and 3GCs-susceptible recipient N. meningitidis Nm040 (penicillin MIC 0.032 µg/mL, cefotaxime MIC 0.008 µg/mL). All natural transformants showed penicillin nonsusceptibility (MIC 0.19–0.38 µg/mL) and cefotaxime resistance (MIC 0.25–0.5 µg/mL). In addition, MICs were elevated by 32–62-fold (0.064–0.125 µg/mL) for ceftriaxone and by 2–5-fold (0.023–0.064 µg/mL) for meropenem compared with recipient Nm040 (ceftriaxone MIC ≤0.002 µg/mL, meropenem MIC 0.012 µg/mL) (Appendix Table 1).
Figure 2
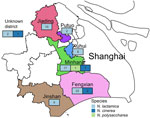
Figure 2. Sampling locations of 76 penA795-bearing Neisseria commensals in 6 districts of Shanghai, China, in study of cefotaxime-resistant Neisseria meningitidis sequence type 4821 causing fulminant meningitis….
Figure 3

Figure 3. Timeline of 85 penA795-bearing Neisseria isolates from China and elsewhere during 2009–2022 in study of cefotaxime-resistant Neisseria meningitidissequence type 4821 causing fulminant meningitis. WHO,…
We cataloged the penA alleles in 1,032 local commensal Neisseria isolates (commensals) and observed penA795 in 76/1,032 (7.4%) isolates. The 76 penA795-bearing commensals included N. lactamica (n = 71), N. cinerea (n = 4), and N. polysaccharea (n = 1) (Figure 1). Among the 71 N. lactamica isolates, CC640 dominated (54.9%, 39 isolates; 27 isolates were ST14031). Those penA795-bearing commensals were collected from 6 districts of Shanghai (Figures 1, 2); they were isolated from persons of various ages, mainly children 3–6 years of age (n = 42), students 7–11 years of age (n = 18), and children <3 years of age (n = 10), over a 10-year period (2013–2022) (Figures 1, 3). All 76 penA795-bearing commensals were resistant to penicillin, cefotaxime, and ciprofloxacin and nonsusceptible (75/76 resistant, 1 intermediate) to trimethoprim/sulfamethoxazole but susceptible to meropenem. Seventy-one isolates (93%, 71/76) were resistant to ceftriaxone (Figure 1; Appendix Table 2).
We identified the penA795 allele among 6 Neisseria species (85 isolates, including 76 from this study) through searches in PubMLST (https://pubmlst.org/neisseria) and PubMed (8,11–14). Of note, penA795 has been transferred among unrelated N. gonorrhoeae clones such as H041 (World Health Organization gonococcal reference strain X [WHO X], 2009, Japan) (11), A8806 (WHO Z, 2013, Australia) (12), GU140106 (2014, Japan) (13), the FC428 international clone (WHO R, 2015, Japan) (8), and G7944 (WHO Q, 2018, United Kingdom) (14) (Figures 1, 3; Appendix Table 1). Only 3 penA795-bearing N. meningitidis isolates have been reported: Nm507 from Shanghai in 2019 and LN24 and LN25 (PubMLST identifications 133563 and 133564) from Shandong Province, China, in 2022. LN24 and LN25 exhibit identical molecular characteristics as B: P1.20,23–1: F1–91:ST5664 (CC4821, L44.2) (Figures 1, 3). MIC data for LN24 and LN25 are unavailable. We extracted the full-length penA (NEIS1753) sequences from 85 penA795-bearing Neisseria isolates and divided them at nucleotide position 718, according to the N terminal (penA 1–717) and C terminal (penA 718–1,746) encompassing the 3 conserved motifs of the PBP2-TPase domain of PBP2 (Appendix Figures 2, 3) (6). The +718 to +900 region possessed 1%–28% nucleotide variations. Except for H041, which had an amino acid alternation of V316P, all isolates harbored the 12 resistance-associated mutations in PBP2-TPase described in Nm507 and FC428 (Appendix Table 1, Figure 3). We classified 85 penA795-bearing Neisseria isolates into 7 groups (a–g) and 6 singletons based on PBP2-TPase sequences associated with cefotaxime resistance (penA 901–1,746, covering 12 resistance-associated mutations) (Figure 1). Nm507 shared an identical cefotaxime-resistant PBP2-TPase sequence with 50 penA795-bearing commensals, as well as LN24, LN25, and A8806 (CTX-R group a), suggesting likely horizontal gene transfer among different Neisseria species, especially from N. lactamica to N. meningitidis. Natural transformation experiments (3) reproduced the horizontal gene transfer event of N. meningitidis acquiring the penA795-bearing segment from Neisseria commensals.
Commensal Neisseria, with a 100% carriage rate, serve as an ideal reservoir of antimicrobial resistance genetic elements for local meningococci. The widespread presence of penA795-bearing N. lactamica CC640, exhibiting multidrug resistance to penicillin, cefotaxime, ceftriaxone, ciprofloxacin, and trimethoprim-sulfamethoxazole in Shanghai, raises substantial concerns over its role in fostering the emergence of multidrug-resistant N. meningitidis. Moreover, the increase of international travel heightens the risk for both meningococcal and multidrug-resistant commensal colonization. Of note, penA795-bearing N. meningitidis (LN24 and LN25) in Shandong were recovered sharing identical PBP2-TPase sequences associated with cefotaxime resistance with commensals from Shanghai.
Our study reveals widespread presence of the penA795 allele, which encodes PBP2-TPase associated with cefotaxime resistance, among Neisseria commensals in Shanghai, China. The penA795 fragment has been captured by the hyperinvasive, quinolone-resistant NmC N. meningitidis ST4821, causing life-threatening meningitis. In China, scheduled meningococcal vaccines include the group A polysaccharide vaccine (MPV-A, 2 doses at 6 and 9 months) and the bivalent NmA and NmC polysaccharide vaccine (MPV-AC, administered at 3 and 6 years of age). This patient had received only 2 doses of MPV-A, because MPV-AC is not licensed for young children (<2–3 years of age). A recent study has just reported an outbreak of NmC caused by multidrug-resistant ST4821 isolates in Fiji (15). Promoting coverage with meningococcal polysaccharide conjugate vaccine (MPCV), such as MPCV-AC or MPCV-ACYW, will be imperative to effectively reduce illness and deaths caused by N. meningitidis, particularly the emerging multidrug-resistant invasive NmC clone.
Dr. Shao is a PhD candidate from Institute of Antibiotics, Huashan Hospital, Fudan University. His research interests include bacterial antimicrobial resistance mechanisms.
[ad_2]
Source link